
Apresentaçőes
2013 - Cromossomos meióticos e mecanismo de determinaçăo do sexo em uma espécie nova de grilo Eneoptera do Nordeste do Brasil (Orthoptera, Eneoterinae) - XXII Congresso de Iniciaçăo Científica da UFPe
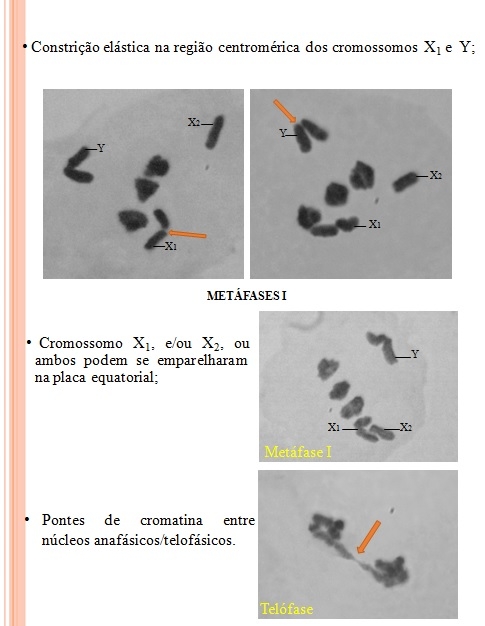
O objetivo deste trabalho foi caracterizar os cromossomos meióticos de uma nova espécie de Eneoptera que ocorre na Floresta Amazônica e Mata Atlântica, destacando o mecanismo de determinação do sexo, bem como o comportamento dos cromossomos sexuais e autossomos durante a Meiose I. Os machos de Eneoptera sp. n. apresentam 2n=9, com sistema múltiplo de determinação sexual X1X2Y. Sendo os autossomos e os cromossomos sexuais X1 e Y metacêntricos e o cromossomo X2 acrocêntrico. Os cromossomos sexuais X1 e Y são heteropicnóticos positivos em relação aos autossomos, desde a fase de leptóteno até diplóteno intermediário. O cromossomo X1 apresenta-se em forma de anel nas fases de zigóteno a paquíteno e o cromossomo Y se apresenta de forma linear ou anelar, com blocos heteropicnóticos positivos distribuídos irregularmente ao longo de toda sua extensão. A heteropicnose positiva dos cromossomos X1 e Y desaparece gradativamente quando os cromossomos avançam para a Metáfase I. A partir dessa fase, os cromossomos sexuais X1 e Y apresentam o mesmo grau de condensação dos autossomos e do cromossomo X2. O cromossomo X2 se comporta como os autossomos, ou seja, é isopicnótico durante toda a meiose I. Durante a metáfase I, torna-se evidente tanto no cromossomo X1 como no Y uma constrição elástica na região centromérica, que pode ser observada de acordo com o seu grau de distendimento do cromossomo. Em algumas metáfases I o cromossomo Y e/ou X1, e/ou X2, ou todos se emparelharam junto aos autossomos na placa equatorial. E em duas anáfases e duas telófases se constatou pontes de cromatina entre núcleos anafásicos. Tanto Eneoptera sp. n. como E. surinamensis apresentam a mesma constituição cariotípica e mecanismos de determinação de sexo, com os machos apresentando 2n=9, X1X2Y. Provavelmente dois passos foram necessários para a origem desse sistema. No primeiro, o cromossomo X metacêntrico ancestral sofreu uma translocação ou fusão com um cromossomo de um par autossômico acrocêntrico, resultando em um metacêntrico grande (Y), o X original remanescente da quebra passou a ser o X1 (acrocêntrico), e o remanescente autossômico, o X2 (acrocêntrico). No segundo passo, houve fusão cêntrica entre o cromossomo acrocêntrico X1 e um cromossomo autossomo acrocêntrico, resultando em um cromossomo X1 metacêntrico (Claus, 1956; Mesa e Bran, 1964). Nos Orthoptera, o comportamento dos cromossomos sexuais se diferencia dos autossomos em relação à condensação cromossômica (Saez, 1963). Após rearranjos entre o cromossomo X com autossomos, os seguimentos de origem autossômica passam gradativamente a se comportar como os cromossomos sexuais. Essa mudança de comportamento pode ser quantificada pelo grau de heterocromatinização e heteropicnose ao longo dos segmentos de origem autossômica (Saez, 1963, Mesa et. al 2001). Embora Eneoptera sp. e E. surinamensis provavelmente tenham herdado o mecanismo de determinação do sexo de um ancestral comum, o cromossomo Y de E. surinamensis apresenta dois blocos heterocromáticos distintos tomando quase toda a extensão de cada braço cromossômico (Piza, 1946; Claus, 1956; Mesa & Bran, 1964) enquanto na espécie aqui analisada, os blocos se distribuem irregularmente ao longo de toda a extensão cromossômica. Outra diferença marcante são as constrições elásticas na região centromérica dos cromossomos X1 e Y, que não ocorrem em E. surinamensis. As pontes anafásicas foram pouco documentadas em Orthoptera, sendo reportadas apenas nos tetrigídeos Tetrix tenuicornis que habitam áreas poluídas com metais pesados (Warcha1owska-Sliwa et. al., 2005). As pontes anafásicas podem ser conseqüência de mutagênicos, os quais aumentam as quebras cromossômicas, com subseqüente fusão de cromátides em ordem invertida, ou promovem a fusão entre telômeros. Os espécimes estudados nesse trabalho foram coletados em uma reserva natural, onde supõe-se que seja reduzida a taxa de poluentes. Novos estudos serão necessários para compreender a origem e o significado dessa anomalia meiótica. Claus, G. 1956. La formule chromosomique du Gryllodea Eneoptera surinamensis De Geer et le comportement des chromosomes sexuels de cette espèce au cours de la spermatogenèse. Ann. Sc. Nat. Paris Zool. 11 (18): 63-106. Hewitt, G.M. Animal citogenetics: Orthoptera (Grasshoppers and crickets). Stuttgart: Gerbruder Borntraeger, 1979, v.3, 170p. (Insecta 1); Mesa, A.; Bran, E.J. 1964. Acerca de los cromosomas de Eneoptera surinamensis. Anais do II Congresso Latino-Americano de Zoologia, São Paulo, 1: 9-16.; Mesa, A.; Fontanetti, C.S.; García-Novo, P. 2001. Does an x-autosome centric fusion in Acridoidea condemn the species to extinction? Journal of Orthoptera Research. 10 (2): 141-146.; Miyoshi, A.R.; Zefa, E., Martins, L.P., Dias, P.G.B.S. Drehmer, C.J., Dornelles, J.E.F. 2007. Stridulatory file and calling song of two populations of the tropical bush cricket Eneoptera surinamensis (Orthoptera, Gryllidae, Eneopterinae). Iheringia, Sér. Zool., 97 (4): 461-465.; Piza Jr, S.T. 1946. Uma nova modalidade de sexo-determinação no grilo Sul-Americano Eneoptera surinamensis. Na. Esc. Sup. Agric. “Luiz de Queiroz” Piracicaba, Brasil, 3: 69-88.; Robillard, T., Desutter-Grandcolas, L. 2005. A revision of Neotropical Eneopterinae crickets (Orthoptera, Grylloidea, Eneopteridae) with a phylogenetic discussion. Insect Systematics and Evolution 35 (4): 411-435.; Saez, F.A. 1963. Gradient of the heterochromatinization in the evolution of the sexual system Neo-X-Neo-Y. Portugaliae Acta Biologica, série A, 7 (1-2): 111-138.; Warcha1owska-Sliwaa, E.; Niklinskab, M.; Gorlichc, A.; Michailovad P.; Pyzac, E. 2005. Heavy metal accumulation, heat shock protein expression and cytogenetic changes in Tetrix tenuicornis (L.) (Tetrigidae, Orthoptera) from polluted areas. Environmental Pollution 133: 373–381. White, M.J.D. Animal citology and evolution. 3.ed. London: Cambridge University Press, 1973. 961p.